|
|
pour en savoir plus :
C. Dammond & D. Tritsch. Neurobiologie
cellulaire, Doin, Paris, 1990.
pour en savoir beauxcoup plus :
L. J. DeFelice. Electrical properties of cells,
Plenum Press, New York, 1997.
|
|
cellule " attachée " (cell-attached patch)
Dans cette configuration, on mesure le courant entre la pipette et lélectrode de résistance. Les variations de résistance correspondent à louverture des canaux " patchés "*. Si un seul canal est ouvert à la fois, on enregistre alors un courant unitaire.
inconvénient : la composition ionique de la cellule et
le potentiel transmembranaire sont mal définis. En effet, la valeur
du potentiel dépend de Vi (potentiel intracellulaire), qui nest
pas connu avec précision. Sur une cellule suffisamment grande (ex:
oocyte de Xenopes), on peut éventuellement imposer un potentiel
à laide de deux électrodes (en plus de lélectrode
de patch). On peut également dépolariser la cellule en utilisant
une solution extracellulaire hyperpotassique.
cellule entière (whole cell clamp)
Le milieu intrapipette dialyse rapidement lintérieur de la cellule, le volume de la pipette étant de beaucoup supérieur à celui de la cellule ; les milieux ioniques sont donc connus. Le potentiel membranaire est connu. Le courant mesuré est un courant global, somme de ceux qui passent par tous les canaux ouverts de la cellule. On peut enregistrer le courant global dun seul type de canaux, en bloquant louverture des canaux dautres types.
inconvénient : on nenregistre pas de courant unitaire
; on ne peut donc pas déterminer la conductance du canal étudié.
La dialyse du milieu intracellulaire entraîne une perte progressive
des constituants intracellulaires.
On étudie le ou les canaux présents dans un fragment de membrane séparé de la cellule. On peut étudier la fonctionnement d'un canal unique.
inconvénient : ne permet pas détudier les canaux
dont le fonctionnement nécessite la présence de constituants
intracellulaires.
configuration outside-out : la face extracellulaire est située à lextérieur de la pipette. On lobtient à partir de la configuration cellule entière. Cest la plus difficile à obtenir. On lutilise lorsquon veut changer la composition du milieu extracellulaire (= extrapipette)
|
|
Quest-ce qui permet de dire que le canal
étudié est un canal dépendant du potentiel (voltage-dependent)
?
Plus on dépolarise, plus le canal est ouvert souvent. À 135 mV, le canal est quasiment toujours fermé (on observe sur la trace uniquement deux évènements douverture du canal, très brefs) ; à 55 mV, il est presque toujours ouvert.
Dautre part, on observe, lorsque que le canal est ouvert, une diminution damplitude du courant unitaire. Cela est dû au fait que plus on dépolarise, plus le potentiel de membrane (Vm) se rapproche du potentiel déquilibre de lion responsable du courant. Ceci nest pas spécifique des canaux dépendants du potentiel (= dont louverture dépend du potentiel de membrane) et sexplique par la loi dOhm.
|
|
En configuration inside-out, le milieu extrapipette correspond au milieu intracellulaire, la partie extrapipette au milieu intracellulaire. Les concentrations ioniques sont proches des concentrations physiologiques.
Composition moyenne des milieux extra- et intracellulaire de Vertébrés
:
| Milieu | Na+ | K+ | Cl- | Ca2+ |
| Extracellulaire (mM) | 14 | 3 | 150 | 1 |
| Intracelllaire (mM) | 14 | 160 | 14 | 10-4 |
calculer les potentiels déquilibre pour chaque ion.
On utilise léquation de Nernst, qui définit la différence de potentiel pour laquelle les gradients électrique et chimique sannulent :
Ei Ee = - (RT/zF)ln ([Xz]i/[Xz]e)
pour une température de 29°C,
Ei Ee = - (60/z)log ([Xz]i/[Xz]e) (mV).
Doù :
ENa = 60 mM EK = -87 mV
réalisation de sauts de potentiel damplitude et de durée variables, à partir dun potentiel de maintien (VH = holding potential)
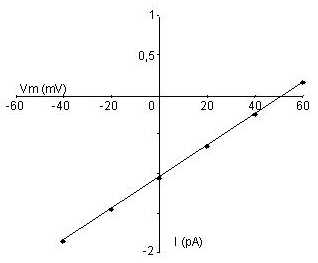
Tracer la courbe i = f(V).
Le potentiel dinversion est de 53 mV. Il est proche du potentiel dinversion de lion Na+ ; le courant est donc principalement dû au mouvement dion Na+. Le canal étudié est un canal sodique.
À partir de la courbe iV, déterminer la conductance (gNa) du canal étudié.
Lintensité du courant dépend de la conductance du canal
et de la force électrochimique appliquée selon la loi dOhm
U = R x I
(la conductance g, exprimée en Siemens
(S) étant égale à linverse de la résistance
R).
La force électromotrice correspond à la différence entre le potentiel membranaire Em et le potentiel d'inversion Einv.
g (Em - Einv) = i = g Em - g Einv
g correspond à la pente de la droite i= f(V) ici, g = 20 pS.
|
|
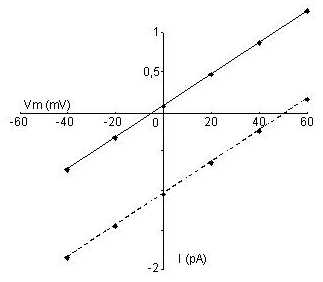
Les concentrations intra- et extrapipette en Na+ étant égales, la droite iV doit passer par 0, si le canal ne laisse passer que le sodium. Or, la droite ne passe pas par 0 et est légèrement décalée vers EK ; le canal est donc légèrement perméable au K+.
Calculer le rapport des conductances au Na+ et au K+ (gNa/gK).
Le courant total passant par le canal est égal à la somme des courants sodique et potassique.
i = iNa
+ iK = gNa(Vm
ENa)
+ gK(Vm
EK)
Au potentiel dinversion Vm, i = o. Doù :
gNa(ENa - Vinv) = gK(Vinv-EK)
gNa / gK = (Vinv-EK) / (ENa - Vinv)
gNa / gK = 19
|
|
Sur les 3 tracés de la figure A, en absence de pronase, on voit quaprès sur phase douverture du canal, il reste à létat fermé alors que la dépolarisation est maintenue. Cela correspond à un état inactivé, au cours duquel le canal ne se rouvre pas malgré le maintien de la dépolarisation.
Sur les traces B, on voit que la pronase ne modifie pas lamplitude du courant unitaire. Par contre, le canal garde sa capacité à souvrir sur toute la durée de la dépolarisation. La pronase supprime donc la capacité du canal à sinactiver.
Pourquoi détermine-t-on la probabilité douverture à létat stationnaire en présence de pronase ?
La probabilité douverture du canal po correspond aux nombre de canaux ouverts sur le nombre de canaux à létat ouvert et à létat fermé.
po = [O]/([O] + [F])
En pratique, on détermine la probabilité douverture du canal en mesurant la durée cumulée durant laquelle le canal est ouvert sur la durée totale de la dépolarisation. Cela suppose que le canal ne puisse être que dans létat ouvert ou fermé, et non dans un état inactivé.
Pour les canaux dépendants du potentiel, comme le courant Na+ étudié, la courbe po = f(Vm) décrit une sigmoïde. Cette courbe sigmoïde définit les caractéristiques de la dépendance vis-à-vis du potentiel.
Il est à noter que les canaux cationiques dont l'ouverture dépend du potentiel sont évolutivement et structurellement reliés. Un canal est formé de quatre sous-unités comprenant chacune 6 domaines transmembraires.

Les propriétés d'inactivation dépendent de la présence d'un domaine de 19 acides aminés sur l'extrémité amino-terminale de chaque sous-unité.
Pour en savoir plus : Molecular Biology of the Cell. Alberts et al, Garlang Publishing Inc. Chap. 11
disponible à la BU de l'Université en traduction française
accessible par chapitre sur PubMed (ouverture dans une nouvelle fenêtre)
|
|
Le courant global sexplique par la sommation des courants individuels. Lensemble des canaux ne souvrent pas et ne se ferment pas en même temps, ce qui explique laspect " lissé " de la courbe globale par rapport à laspect " en carré " des courants unitaires. Le point damplitude maximale du courant correspond au moment où le plus grand nombre de canaux sont dans létat ouvert. Le retour progressif à un courant nul est dû à linactivation progressive des canaux.
Pourquoi les expériences ont-elles été effectuées en présence de TEA ?
Lenregistrement du courant global correspond à lenregistrement de lensemble des courants passant par les différents types de canaux ioniques, principalement les courants de sodium et de potassium. Le TEA bloque les canaux K+, ce qui permet de nenregistrer que le courant Na+.
|
|
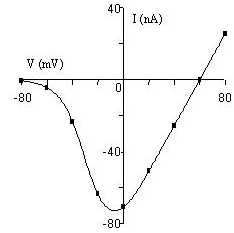
Déterminer le potentiel dinversion. Le comparer au potentiel déquilibre des ions Na+ et K+. Que peut-on en déduire sur la nature du courant ?
Le potentiel dinversion est égal à 60 mV.
Les potentiels déquilibre des ions Na+ et K+ sont les suivants, déterminés par léquation de Nernst :
ENa = 60 mM EK = -87 mV
Le potentiel dinversion Vm est égal au potentiel déquilibre du Na+. Il sagit donc dun courant de sodium.
|
|
Le récepteur nicotinique est un exemple-type de ce types de canaux,
dont l'ouverture dépend de la liaison à un agoniste. Les
principaux canaux de ce type sont donnés dans le tableau suivant
:
| type de canal | effet |
| canaux cationiques activés par l'ACh | excitateur |
| canaux cationiques activés par la 5-HT | excitateur |
| canaux cationiques activés par le glutamate | excitateur |
| canaux Cl- activés par le GABA | inhibiteur |
| canaux Cl- activés par la glycine | inhibiteur |
Les canaux cationiques sont excitateurs car leur ouverture entraîne
un influx de cations et donc une dépolarisation, alors que l'activation
des canaux Cl- s'oppose à la dépolarisation.
Quel serait laspect dune courbe IV dun canal dépendant dun agoniste, ou mécano sensible ?
Le courant global I est le produit du courant unitaire i, du nombre de canaux N et de la probabilité douverture po. I = i.N.po
N est une constance ; i dépend du potentiel de manière linéaire.
Pour un canal dépendant du potentiel, po décrit, en fonction du potentiel, une sigmoïde. Laspect de la courbe I(courant global)V résulte de la conjonction de ses deux paramètres, la courbe sigmoïde de po et la droite de i.
Pour un canal non dépendant du potentiel, po de dépend pas de V. En absence de stimulation (agoniste, variation de pression ), le courant sera nul, quel que soit le potentiel. En présence dune stimulation, po prendra une valeur donnée, dépendante de lintensité de la stimulation, mais indépendante du potentiel. La courbe IV sera alors une droite.
modélisation du fonctionnement du canal Cl- dépendant du Ca2+ dans les myocytes des voies aériennes.
Comment varie lintensité du courant en fonction de la concentration cytosolique en calcium ?
L'intensité est nulle lorsque la concentration cytosolique en calcium est proche de la valeur de repos (100 nM), et elle augmente de manière sigmoïdale en fonction de l'élévation de la concentration en Ca2+.
Comment varie lintensité du courant en fonction du potentiel
de membrane ? pourquoi ?
L'intensité du courant, pour une valeur de concentration en
calcium donnée, varie linéairement en fonction du potentiel.
Cela signifie que la probabilité d'ouverture ne dépend pas
du potentiel.
Est-il intéressant de faire des courbes IV sur des canaux indépendants du potentiel ?
Lintérêt des courbes IV sur des courants passant par des canaux indépendants du potentiel est de caractériser ces courants, par comparaison du potentiel dinversion et du potentiel déquilibre des ions éventuellement responsables du courant.
| Etienne Roux UFR SV UB2 |
|