
classification fonctionnelle :

|
|
|
|
Comment peut-on classer les différents types de muscles ?
On peut caractériser un muscle, en tenant compte :
1. de l'organisation interne de la cellule musculaire
On peut distinguer d'une part les muscles squelettiques et cardiaques, d'autre part les muscles lisses. Dans les muscles squelettiques et cardiaques, les myofilaments sont organisés en sarcomères. Ce sont des muscles à contraction rapide et de relativement courte durée. Dans les muscles lisses, les myofilaments ne sont pas organisés en sarcomères. Ce sont des muscles à contraction lente et maintenue.2. de l'organisation du tissu musculaire
On peut opérer un autre type de classement, en distinguant d'une part les muscles squelettiques, qui agissent par effet de levier, et d'autre part les muscles cardiaques et lisses, qui se trouvent dans la paroi d'organes creux.3. du contrôle de l'activité musculaire par le système nerveux
On peut ainsi distinguer les muscles volontaires des muscles involontaires, bien que cette terminologie ne soit pas très appropriée, et opérationnelle uniquement chez lhomme. Il est préférable de distinguer les muscle sous contrôle du système nerveux moteur et ceux sous contrôle du système nerveux autonome.En fonction de ces critères, on peut grouper les muscle de différentes façons :
Comment peut-on classer les différents types de muscles
?
Si lon privilégie laspect anatomique ou laspect fonctionnel,
on aboutit à deux classifications des muscles. La classification
anatomo-histologique regroupe les muscles squelettiques et cardiaques,
distingués des muscles lisses, alors qu'une classification fonctionnelle
peut regrouper les muscles cardiaques et lisses présents dans
la paroi d'organes creux, et contrôlés par le système
nerveux autonome distingués des muscles squelettiques.
classification anatomique :
classification fonctionnelle :
|
|
Quelles sont les caractéristiques communes aux muscles
squelettique et cardiaque ?
L'organisation du système contractile en monomère et
la présence de myoglobine, associées à la capacité
de contraction rapide et de courte durée.
Quelles sont les caractéristiques communes aux muscles lisse
et cardiaque ?
La présence du muscle dans la paroi d'organes creux ; la présence
de myocytes monocucléés ; l'existence de jonction gap, qui
sont des jonctions électriques de cellule à cellule ; une
régulation complexe de la contraction, impliquant entre autre le
système nerveux autonome et une régulation hormonale.
NB : il existe plusieurs types de muscle lisse. On distingue :
les muscles lisses mono-unitaires
ils sont caractérisés par la présence de jonctions
de cellules à cellules, et la transmission d'un potentiel d'action
de cellule à cellules. Ils sont caractérisés par un
type de contraction dit phasique.
ex : myocytes responsables du péristaltisme intestinal.
les muscle lisses pluri-unitaires
il n'y a pas dans ces myocytes de potentiel d'action, et pas de propagation
électrique de cellule à cellule. Ils sont caractérisés
par une contraction dite tonique.
ex : les myocytes des voies aériennes, responsables de la
réactivité bronchique.
|
|

Le schéma représente deux sarcomères. Lorganisation en sarcomères est caractéristique du muscle strié. Les fibres musculaires striées squelettiques sont constituées de grandes cellules plurinucléées, dont le diamètre est de 0,01 à 0,1 mm, et la longueur très variable. L'appareil contractile est organisé sous forme de sarcomères, de 2,5 µm de long, séparés par des stries Z. Dans le muscle cardiaque, l'organisation des myofilaments est à peu près la même, mais le myocyte cardiaque est une cellule mononucléée. La contraction se propage à travers la masse musculaire de cellule à cellule.
La cellule musculaire lisse est différente, tant d'un point de vue microanatomique que d'un point de vue fonctionnel. Les caractéristiques de l'organisation de la cellule musculaire lisse seront présentées plus loin.
Représentez le même schéma, mais lorsque le muscle est à son état de contraction maximum.

Pour
en savoir plus : Molecular Biology of the Cell. Alberts et al,
Garlang Publishing Inc.
disponible à la BU de l'Université
en traduction française
accessible par chapitre sur PubMed
(ouverture dans une nouvelle fenêtre)
|
|
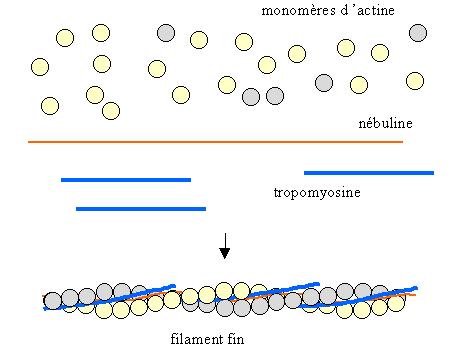
actine : monomères dactine (PM = 43 000) organisés en un filament à deux brins
tropomyosine : molécule composée de deux chaînes polypeptidiques, associée au filament dactine
nébuline : protéine du cytosquelette
|
|
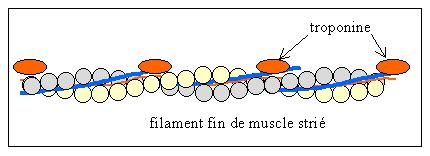
Les monomères d'actine s'organisent en un filament à deux brins autour de la nébuline. Les molécules de tropomyosine viennent se fixer dans le sillon central. Chaque molécule de tropomyosine s'étend sur environ 7 monomères d'actine.
|
|
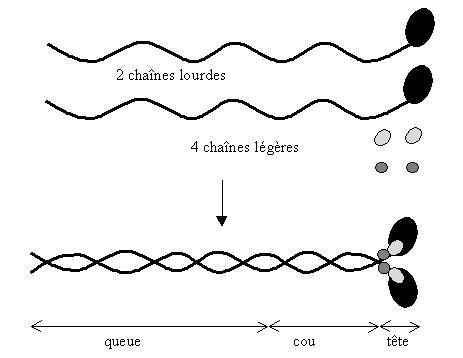
un monomère de myosine est une molécule constituée
de 6 polypeptides, 2 chaînes lourdes et 4 chaînes légères.
Les deux chaînes lourdes sont organisées en hélices
alpha, enroulées en superhélice.
Les chaînes légères sont responsables de l'activité
ATPasique. De cette activité ATPasique dépend la vitesse
du cycle de contraction ainsi que la consommation d'ATP.
Un monomère de myosine mesure environ 150 nm. On distingue trois
parties: la queue (93 nm), le cou (57 nm) et la zone globulaire qui constitue
la tête.
|
|
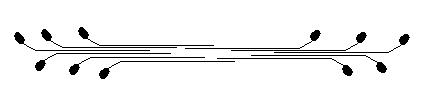
Les filaments de myosine sont constitués de l'agrégation d'un nombre variable de monomères. Cette agrégation peut être spontanée (en fonction de la forme ionique) et aboutit à la constitution de filaments de 1,5 µm de long et 12 nm de diamètre.
Dans une cellule, le filament épais est constitué de 20 à 400 molécules de myosine, selon le type cellulaire.
Les muscles striés et les muscles lisses diffèrent quant
à leurs isomères de myosine.
De même, il existe plusieurs types de fibres musculaires striées,
en fonction des isoformes de myosine qu'elles contiennent. Chez l'homme
adulte, il existe 3 isoformes de la chaîne lourde myosine : I, IIa
et IIx, contenues dans 3 differents types de fibres, du même nom.
Les fibres de type I sont dites "lentes", et les fibres de type IIa et
IIx sont dites "rapides". La vitesse de contraction d'une fibre de type
I est dix fois plus faible que la vitesse de contraction d'une fibre de
type IIx.
Pour en savoir
plus :
Structure et fonctionnement du muscle :
Molecular Biology
of the Cell. Alberts et al, Garlang Publishing Inc.
disponible à la BU de l'Université
en traduction française PubMed
(ouverture dans une nouvelle fenêtre)
Physiology,
Berne et al, Mosby, Saint-Louis (4th edition, 1998). (voir
section III Muscle)
rappels sur la contraction musculaire et les
différents types de fibres musculaires :
Andersen et al,
Muscle, gènes et performances (2000) Pour la Science, 276: 32-41.
(disponible BU)
|
|
expérience n° 2: (ouverture dans une nouvelle fenêtre)
Que peut-on déduire des deux expériences précédentes du rôle du calcium et de l'ATP dans la contraction du muscle strié?
On sait que la contraction musculaire nécessite de l'énergie, fournie par l'hydrolyse d'ATP. Toutefois, les résultats montrent que la seule présence d'ATP ne suffit pas à engendrer la contraction musculaire. La présence de calcium est également nécessaire.
La présence simultanée de calcium et dATP nécessaire au développement de la force contractile. Par contre, si la suppression du calcium entraîne une relaxation, le maintien de la contraction en présence d'ATP est possible en absence de calcium.
|
|
|
|
1. En absence de calcium, la troponine masque le site de fixation de la tête de myosine sur l'actine. Il n'y a pas de formation du pont actomyosique, y compris en présence d'ATP. La myosine a une forte affinité pour l'ADP et le phosphate inorganique (Pi), qui sont fixés sur la tête de myosine. Lorsque l'ADP et Pi sont fixés à la tête de myosine, la conformation pour laquelle l'energie libre est minimale lorsque la tête de myosine fait un angle de 90° avec le filament d'actine.
2. En présence de calcium, qui se fixe sur la sous-unité C de la tropomyosine, celle-ci démasque le site de fixation de la myosine sur l'actine. Il y a formation du complexe actomyosique.
3. Lorsque la myosine est fixée sur l'actine, son affinité pour l'ADP et Pi diminue. Ceux-ci se détachent de la tête de myosine.
4. En absence d'ADP et de Pi, la conformation pour laquelle l'énergie libre du complexe actomyosique est la la plus faible correspond à un angle de 45° entre la tête de myosine et le filament d'actine. Il y a donc rotation de la tête de myosine, diminution de l'énergie libre du complexe actomyosique, et génération d'une force contractile (correspondant à cette variation d'énergie libre). La rotation de al tête de myosine entraîne un déplacement de 10 nm du filament d'actine par rapport au filament de myosine.
5. Dans cette conformation, la tête de myosine a une forte affinité pour l'ATP, qui vient s'y fixer.
6. L'activité ATPasique de la tête de myosine entraîne une hydrolyse de l'ATP en ADP et Pi, et la rupture du pont actomyosique. Lorsque la tête de myosine est séparée de l'actine et liée à l'ADP et au Pi, sa conformation correspondant à l'energie libre minimale correspond à un angle de 90° entre l'actine et la tête de myosine.
a) En absence de calcium, le cycle s'arrête en position 1.Pour en savoir plus :
b) En présence de calcium, le cycle repart à partir de la position 2.
|
|
Expliquer les différentes phases de lexpérience
présentée.
(les numéros font références aux différentes étapes commentées dans l'exercice précédent)
En présence de calcium, le complexe actomyosique se forme, et un premier cycle de contraction a lieu. Mais en absence d'ATP, il s'arrête en position 4. Il n'y a pas développement de la contraction musculaire.
Lorsqu'on ajoute du calcium, le cycle peut s'effectuer, et il y a génération de la contraction. La tension augmente jusqu'à une valeur plateau.
Lorsqu'on enlève l'ATP, le cycle s'arrête en position 4, alors que les ponts actomyosiques sont constitués. Le cycle de contraction est arrêté, mais le maintien des ponts actomyosiques fait que la tension précédemment engendrée se maintient également (sans consommation d'ATP). C'est ce qui se passe dans le phénomène de rigidité cadavérique. Après la mort, l'arrêt des pompes calciques dû à l'absence d'ATPqui n'est plus produit entraîne une augmentation de la concentration cytosolique en calcium. Ceci entraîne la formation des ponts actomyosiques, responsables de la rigidité cadavérique.
Pourquoi, en présence de calcium, la contraction se maintient-elle
en absence dATP ?
La situation précédente (arrêt du cycle en position
4) se maintient également en absence de calcium.
Pourquoi, en absence de calcium, lajout dATP provoque une relaxation
? Quobserverait-on en présence de
calcium ?
L'ajout d'ATP provoque la rupture des ponts actomyosiques, qui, en
absence de calcium, ne peuvent se reformer. Il y a donc relaxation. En
présence de calcium, le cycle de contraction reprendrait et une
tension serait générée (avec consommation d'ATP).
Force contractile développée par une fibre musculaire perméabilisée en fonction de la concentration du milieu en calcium
Comment peut-on expliquer l'aspect de la courbe reliant la force développée par une fibre musculaire et la concentration intracellulaire en calcium?
Lorsquune cellule est perméabilisée, le calcium du milieu pénètre dans la cellule et la concentration cytosolique devient alors égale à celle du milieu. La courbe expérimentale représente donc, en coordonnées semi-logarithmique, lintensité de la contraction en fonction de la concentration cytosolique en calcium. La dépendance de la contraction vis-à-vis du calcium et laspect sigmoïde de la courbe sexpliquent de la manière suivante :
En absence de calcium, la tropomyosine masque la fixation de la myosine sur l'actine. En présence de calcium, qui se fixe sur la sous-unité C de la tropomyosine et entraîne un changement conformationnel de celle-ci, le site de fixation de la myosine sur l'actine est déplacé, ce qui permet la constitution du complexe acto-myosique. Selon ce mécanisme, la formation de ponts actomyosiques, responsables de la contraction et donc lintensité de la contraction dépend de la concentration en calcium cytosolique. Ceci explique laspect sigmoïde de la courbe. Cet aspect correspond à une courbe de liaison ligand-récepteur, la liaison étant ici celle du calcium avec la troponine C.
On peut noter que l'intensité de la contraction dépend de la concentration en calcium dans une gamme de concentration qui correspond à la gamme de variations de la concentration cytosolique en calcium d'une cellule.
|
|
Pourquoi l'étirement passif du muscle nécessite-t-il
une tension?
Parce que le muscle est élastique. Lélasticité
dun myocyte isolé est due à lélasticité du
cytosquelette. Lélasticité dun muscle est plus importante,
car sy ajoute lélasticité du tissu conjontif qui entoure
les faisceaux musculaires.
La force nécessaire pour étirer le muscle relâché
est appelé la force passive ou tension passive.
NB: En biophysique musculaire, on fait la distinction entre la force
(force) et la tension (stress), la tension étant la force par surface
de section trasnversale (cross-sectional area).
Comment expliquer l'aspect de la courbe de tension active ? Qu'est-ce
que le L0 ?
Lorsque le muscle est stimulé, il génère une force
ou tension active qui sajoute à la force ou tension passive.
La tension maximale développé par le muscle dépend
de sa longueur initiale avant stimulation. La courbe longueur initiale-tension
active a un aspect en cloche, et la longueur pour laquelle la tension développée
est maximale est appelée L0. La
force active est proportionnelle au nombre de ponts actomyosiques qui interagissent
dans chaque sarcomère dont la longueur est a peu près constante
, et donc au chevauchement des filaments. Lorsque le muscle est étiré
au delà de L0, le chevauchement
des filaments est incomplet. lorsque le muscle est étiré
à 150 % de L0, il ny a plus de
chevauchement et la force générée est nulle.
À linverse, lorsque la longueur est égale à 70
% de L0, les lignes Z ont rejoint les filaments
de myosine, et le muscle ne peut plus se raccourcir. De plus, en deçà
de L0, les éléments élastiques
absorbent une partie de la force générée par lappareil
contractile.
|
|
Que peut-on dire de la relation entre la tension initiale et la vitesse
de raccourcissement ?
Qu'est-ce que V0 ? De quoi dépend-il ?
La vitesse de raccourcissement dépend, pour un muscle donnée, de la tension passive exercée sur le muscle. Le raccourcissement est le plus rapide lorsque la muscle nest pas étiré (V0). Plus la tension exercée devient importante, plus la vitesse diminue, jusquà devenir nulle ; lorsquon augmente encore la tension, la vitesse devient négative, cest-à-dire que le muscle sallonge sous leffet de la tension. Au début, Cet allongement est lent, puis il saccroît rapidement.
V0 est la vitesse maximale de raccourcissement. Etant donné que la vitesse de raccour-cis-sement est fonction du nombre de sarcomères en série, V0 en général est exprimé par longueur de ½ sarcomère. V0 dépend de la vitesse dhydrolyse de lATP par la myosine, et dépend donc de lisoforme de myosine présente dans le muscle. Dans le muscle striée, il existe principalement deux types de fibres, des fibres lentes (type I) et des fibres rapides (type IIx et IIa) correspondant a 3 isoformes de mysosine. Les fibres IIx, les plus rapides, se contractent 10 fois plus vite que les fibres I.
V0 correspond au taux maximum de formation des ponts actomyosiques. Ils se forment très rapidement, et il y a peu de ponts existant à un moment donné. Lorsque la tension augmente, le nombre de ponts présents à un moment donné augmente (la force développée dépend du nombre de ponts actomyosiques formés), ce qui correspond à un taux de formation des ponts actomyosiques plus faible.
Lorsque la tension devient plus forte que celle que le muscle peut développer, le muscle sallonge. Tant que la tension exercée nexcède pas 1,6 fois celle que le muscle peut développer, les ponts actomyosiques persistent. Au-delà, les ponts actomyosiques sont rompus et la vitesse dallongement augmente brusquement.
rappels sur la contraction musculaire et les
différents types de fibres musculaires :
Andersen et al,
Muscle, gènes et performances (2000) Pour la Science, 276: 32-41.
(disponible BU)
|
|
structure
Lappareil contractile, bien que composé de filaments fins et
de filaments épais, nest pas organisé en sarcomères.
Les
filaments fins ne contiennent pas de troponine.
fonctionnement
Lactivité ATPasique de la myosine du muscle lisse, nécessaire,
comme dans le muscle strié, à la contraction musculaire,
nest pas due au démasquage des sites de fixation de lactine sur
la myosine par déplacement de la troponine, mais à la phosphorylation
de la chaîne légère de 20 kDa de la myosine.
Pour en savoir
plus :
Molecular Biology
of the Cell. Alberts et al, Garlang Publishing Inc.
disponible à la BU de l'Université
en traduction française PubMed
(ouverture dans une nouvelle fenêtre)
|
|
|
|
Comment peut-on expliquer l'aspect de la courbe reliant la force développée par un lambeau musculaire lisse et la concentration intracellulaire en calcium? Quelles sont les différences avec un faisceau musculaire strié?
D'après les résultats présentés, le calcium, comme dans la cellule musculaire striée, est nécessaire à la contraction de la cellule musculaire lisse. La génération de la force musculaire dépend de la concentration cytosolique en calcium (dans les conditions expérimentales où les cellules sont perméabilisées, les concentrations extracellulaires et cytosoliques en calcium s'équilibrent).
Toutefois, il existe des différences importantes entre le fonctionnement de l'appareil contractile de la cellule musculaire striée et la cellule musculaire lisse. En particulier, dans la cellule musculaire lisse, il n'existe pas de troponine qui empêche, en absence de calcium, la formation du complexe actomyosique et l'activité ATPasique de la myosine. L'activité ATPasique et la myosine et sa capacité à former un complexe actomyosique nécessite la phosphorylation de la chaîne légère de 20 kD de la myosine. Cette phosphorylation permet la fixation de la myosine sur l'actine, et c'est la formation du complexe actomyosique qui permet l'activité ATPasique de la myosine (cf. schéma) La phosphorylation de la MLC20 est assurée par la kinase de la chaîne légère de la myosine (MLCK), elle-même activée par le complexe calcium-calmoduline, dont la formation dépend de la concentration cytosolique en calcium.
Ceci explique que laspect de la courbe de la force générée en fonction de la concentration en calcium ait le même aspect sigmoïde dans le muscle lisse et dans le muscle strié. Dans le premier, laugmentation en calcium joue sur la concentration en complexe Ca-calmoduline, alors que dans le muscle strié, elle joue sur la concentration en complexe Ca-troponine C.
|
|
À quels niveaux intervient le calcium dans le cycle de contraction
?
A toutes les étapes du cycle de contraction, puisqu'il va jouer,
à chaque étape, sur l'équilibre entre les formes phosphorylées
et non phosphorylées de la myosine (par modulation de la MLCK).
À quels niveaux seffectue la consommation dATP ?
Comme dans le cycle de contraction de la cellule musculaire striée,
une molécule d'ATP sera consommée par cycle de contraction,
hyprolysée grâce à l'activité ATPasique de la
chaîne légère de la myosine ; mais de l'ATP également
sera consommé à chaque phosphorylation de la chaîne
légère de la myosine. Par cycle de contraction l'appareil
contractile de la cellule musculaire lisse consomme donc plus d'ATP que
celui de la cellule musculaire striée.
Lorsque la chaîne légère de la myosine est déphosphorylée,
le cycle seffectue à une vitesse plus faible que lorsquelle est
phosphorylée.
Quel va être la conséquence dune diminution de la proportion
de phosphorylation :
sur la vitesse de raccourcissement ?
En cycle non phosphorylé, le cycle est ralenti. La vitesse de
raccourcissement est donc diminuée.
sur la consommation dATP ?
La consommation d'ATP étant dépendante de la vitesse
du cycle (une molécule d'ATP consommée par cycle), la consommation
d'ATP sera ralentie.
Comment ce système permet-il de maintenir une force importante
avec une faible consommation dénergie ? Est-ce compatible avec
une vitesse de raccourcissement élevée ?
En cycle non phosphorylé, le cycle est ralenti. Le temps durant
lequel le complexe actomyosique est formé est augmenté. Or,
durant ce temps, la contraction est maintenue sans consommation d'ATP.
Ce système permet le maintien mais pas l'établissement
d'une force importante avec une faible consommation d'énergie.
Mais ce n'est pas compatible avec une vitesse de raccourcissement élevée.
Ainsi, la modulation de la vitesse du cycle de contraction par l'existence d'un cycle de contraction partiellement déphosphorylé existe dans le muscle lisse mais pas dans le muscle strié. Ceci permet de découpler d'une part la vitesse de raccourcissement et la génération de la force, et d'autre part le maintien de la tension. Ceci n'est pas possible dans le muscle strié. Le muscle lisse est donc capable de maintenir une contraction avec une faible consommation d'ATP (300 fois inférieure à la consommation en ATP du muscle strié).
NB : lhypothèse des ponts verrouillés
rend compte des observations, mais na pas été formellement
démontrée. En particulier, le pont actomyosique verrouillé
na pas pu être isolé biochimiquement.
Divers mécanismes peuvent expliquer
lexistence de cycles de contraction déphosphorylés. La la
caldesmone (CD) et la calponine(CP), des protéines associées
au filament fin peuvent moduler la contraction. Non phosphorylées,
elles diminuent la contraction ; phyosphorylées, elles l'augmentent.
Des résultats récents suggèrent fortement limplication
de protéine Rho et dune autre isoforme de la myosine, la MLC17,
dans la phase maintenue et déphosphorylée de la contraction.
Pour en savoir
plus :
Physiology,
Berne et al, Mosby, Saint-Louis (4th edition, 1998). (voir
section III Muscle, chap. 19)
Horowitiz et
al, Mechanisms of smooth muscle contraction (1996), Physiol
Rev 76: 967-1003 (disponible BU)
|
|
Sur un lambeau musculaire lisse, on mesure simultanément
la contraction isométrique et la
[Ca2+]i.
L'hypothèse des ponts actomyosiques verrouillés permet-elle
d'expliquer les résultats obtenus ? Quelle autre explication pourrait
rendre compte des observations expérimentales ? Quelle mesure pourrait-on
faire pour choisir entre les deux explications ?
Oui. La phase de développement de la tension,
associée à une [Ca2+]i
élevée, correspond à un cycle de contraction phosphorylée.
La deuxième phase, lorsque la [Ca2+]i
est diminué, correspond à un cycle lent partiellement déphosphorylé
(latch bridge cycle).
Une autre possiblité de régulation pouvant expliquer la figure ci-dessus met en jeu une inhibition de la MLCP. Linhibition de la MLCP augmente, pour une [Ca2+]i donnée, la proprotion de PLC20 phosphorylée ; ceci correspond à une augmentation de la sensibilité de lappareil contractile au calcium, et peut expliquer les résultats obtenus.
Dans la première hypothèse, le taux de phosphorylation de la MLC20 est faible, alors que dans la seconde hypothèse, il demeure élevé malgré la diminution de la [Ca2+]i. Une mesure de la phosphorylation de la myosine permettrait de trancher entre les deux hypothèses.
Courbe tension-vitesse de raccourcissement
Comparez les différentes courbes. Comment
s'explique la faible vitesse de raccourcissement du muscle lisse par rapport
au muscles squelettiques?
Les courbes concernant les muscles striés
correspondent à des muscles qui diffèrent quant aux isoformes
de la myosine. Le msucle lisse se distingue des muscles striés par
une myosine différente dont lactivité ATPasique est caractérisée
par une cinétique beaucoup plus lente. Dautre part, lorganisation
de la cellule musculaire lisse, où il ny a pas de sarcomère,
diffère de celle du muscle strié, ce qui jour également
sur la vitesse de raccourcissement.
Dans le muscle lisse, de quoi dépend
V0 ?
V0 dépend de la cinétique
lactivité ATPasique de la myosine, qui dépend de lisoforme
de myosine présente mais également du taux de phosphorylation
de la myosine.
|
|
différents types de contraction
Donnez la définition de ces différents types de contraction
:
contraction isométrique : contraction qui seffectue
à longueur constante
contraction isotonique : contraction qui seffectue à
charge constante
contraction auxotonique : contraction qui seffectue à
charge et longueur variable
Quel(s) type(s) de contraction corresponde(ent) à un fonctionnement
physiologique ?
Essentiellement une contraction auxotonique, car la charge contre laquelle
s'effectue la contraction musculaire est en général modifiée
lorsque le muscle se raccourcit. Il peut exister aussi une contraction
isométrique, plus rarement isotonique.
relation longueur-tension
On exprime souvent la tension par rapport à la surface de section
transversale du muscle. Pourquoi?
La force dépend du nombre de ponts actomyosiques qui peuvent
se former. La longueur d'un sarcomère (2,5 µm) et la densité
des filaments contractiles étant constantes, la force va varier
en fonction de la surface de section transversale. En déterminant
la tension (force/surface de section transversale) on peut comparer les
forces de plusieurs muscles, indépendamment de la taille du muscle
sur lequel on effectue les mesures de contraction.
Si on réalise un courbe longueur-tension sur un muscle lisse,
on s'aperçoit que l'aspect de la courbe est similaire à celui
de la courbe longueur-tension d'un muscle strié. Que peut-on en
conclure ?
On peut en conclure que, bien que lappareil contractile du muscle
lisse ne soit pas organisé en sarcomère, la force de contraction
est généré par le même type de mécanisme,
le glissement des filaments fins et épais par la formation de ponts
actomyosiques.
vitesse de raccourcissement
On exprime la vitesse de raccourcissement en µm/sec par longueur
de 1/2 sarcomère. Quel est l'intérêt de cette normalisation
? On l'exprime souvent également en L0/sec. Pourquoi ?
Cette normalisation permet de comparer plusieurs muscle entre eux.
Les deux normalisation sont équivalentes, puisque L0 rend
compte du nombre de qarcomètres en série dans un muscle.
force, travail, puissance
Si on met bout à bout deux muscles identiques, quelle sera la force développée par ces deux muscles ensemble par rapport à la force développée par chacun deux séparément ? Quel sera le travail fourni ?
La force est proportionnelle à l'aire de section d'un muscle,
la longueur d'un sarcomère (2,5 µm) et la densité des
filaments contractiles par surface de section musculaire étant constantes.
La force générée par deux muscles identiques mis bout
à bout sera égale à la force générée
par un seul des deux. Le poids maximal soulevé par les deux muscles
est égal au poids maximal d'un seul des deux.
Le travail, lui, est égal au poids de la charge déplacé
par la distance. Pour deux muscles placés en série,
la force est identique mais le déplacement est doublé ; le
travail est donc multiplié par 2.
La force développée par un muscle dépend-elle de sa longueur, de son épaisseur, de son volume ?
La force générée maximale générée par un muscle striée, quelque soit l'espèce (la longueur d'un sarcomère (2,5 µm) et la densité des filaments contractiles par surface de section musculaire étant constantes), est de 39,3 à 49 N/cm2.
NB : il est de très rares cas où la force est modifiée. Les sarcomères du muscle rétracteur de la moule ont une longueur de 25 µm, et la force maximale générée atteint 100 N/cm2.
Quest-ce que la puissance dun muscle ?
La puissance d'un muscle est le travail divisé par le temps, ou la force multipliée par la vitesse.
À force identique, un muscle lisse sera-t-il plus ou moins puissant quun muscle strié ?
La vitesse de contraction d'un muscle lisse étant beaucoup plus faible que celle d'un muscle strié, sa puissance sera également plus faible.
À longueur et épaisseur identiques, un muscle de souris est-il plus ou moins puissant quun muscle déléphant ?
Pour une longueur et une épaisseur identiques, un muscle de souris développe une force et un travail identique à un muscle d'éléphant. Par contre, la vitesse de raccourcissement d'un muscle de souris est plus rapide que celle d'un muscle d'éléphant (en général, les animaux de petite taille ont un métabolisme et une consommation d'ATP plus élevés que les animaux de grande taille).
Pour une longueur et une épaisseur identiques, un muscle de souris
est donc plus puissant qu'un muscle d'éléphant.
| Etienne Roux UFR SV UB2 |
|